3/1/2019
LEDs: Far Red & Light Intensity Interaction
Yujin Park & Erik Runkle
 Indoor plant production enables growers to precisely control environmental conditions such as light, temperature, humidity and CO2 to consistently produce uniform crops. In addition, using light-emitting diodes (LEDs) indoors makes it possible to manipulate the light spectrum to elicit desirable plant attributes, such as compact growth, and early or late flowering.
Indoor plant production enables growers to precisely control environmental conditions such as light, temperature, humidity and CO2 to consistently produce uniform crops. In addition, using light-emitting diodes (LEDs) indoors makes it possible to manipulate the light spectrum to elicit desirable plant attributes, such as compact growth, and early or late flowering.
Most of the plant research with LED lighting has focused on blue (400 to 500 nm) and red (600 to 700 nm) light. Increasing blue light intensity generally promotes plant pigmentation and suppresses extension growth, resulting in more compact plants. The influence of including additional wavebands, such as green (500 to 600 nm) and far-red (700 to 800 nm) light, has been explored more recently. Including far-red light in indoor lighting has demonstrated potential merit for crop production, such as greater leaf expansion and growth, and accelerated flowering in some floriculture crops.
Plant leaves absorb most photons within the photosynthetically active radiation waveband (400 to 700 nm). In contrast, far-red light is poorly absorbed, and most is transmitted through or reflected by leaves. Plants perceive and acclimate to the red to far-red ratio (R:FR). As the R:FR decreases (or as relative portion of far-red light increases), shade-avoidance responses—such as stem elongation, leaf expansion and early flowering—are stimulated.
However, plant responses to far-red light (or the R:FR) can be affected by light intensity (or more technically, the photosynthetic photon flux density, PPFD) and blue light intensity. In addition to a low R:FR, shade-avoidance responses can also be triggered by a low PPFD and/or low blue light. We investigated how PPFD influences the effects of adding far-red light on seedling growth and subsequent flowering in three floriculture crops. (In the next article of this series, we’ll present how far-red light interacts with blue light to regulate growth attributes of young plants.)
Experimental protocol
Seeds of Geranium Pinto Premium Orange Bicolor, Petunia Wave Blue and Coleus Wizard Golden were sown in 128-cell plug trays by Raker-Roberta’s and received at Michigan State University (MSU) about one week later. When the first true leaves emerged, the seedlings were thinned to one plant per cell. They were then grown under six LED lighting treatments (Table 1) in a refrigerated growth chamber at a constant 68F (20C) and an 18-hour photoperiod.
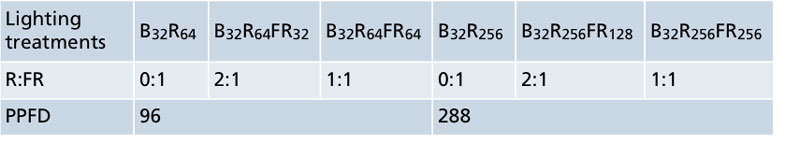 Table 1. The ratio of red (R, 600 to 700 nm) to far-red (FR, 700 to 800 nm) light and photosynthetic photon flux density (PPFD, values in µmol•m–2•s–1) of six indoor lighting treatments. The subscript value after each LED type (B = blue, R = red, FR = far red) indicates its photon flux density in µmol•m–2•s–1.
Table 1. The ratio of red (R, 600 to 700 nm) to far-red (FR, 700 to 800 nm) light and photosynthetic photon flux density (PPFD, values in µmol•m–2•s–1) of six indoor lighting treatments. The subscript value after each LED type (B = blue, R = red, FR = far red) indicates its photon flux density in µmol•m–2•s–1.
The LED lighting treatments were provided by different mixtures of blue, red and/or far-red LEDs to investigate how far-red light influenced growth at two PPFDs, 96 µmol•m–2•s–1 (PPFD 96) and 288 µmol•m–2•s–1 (PPFD 288). Growth attributes of seedlings were evaluated after approximately three to five weeks under the LEDs, when they were ready for transplant.
 Figure 1. Influence of adding far-red light on seedling height (values in cm) of petunia, geranium and coleus. The subscript value after each LED type (B = blue, R = red, FR = far red) indicates its photon flux density in µmol•m–2•s–1. Seedling heights followed by the same letter are statistically similar.
Figure 1. Influence of adding far-red light on seedling height (values in cm) of petunia, geranium and coleus. The subscript value after each LED type (B = blue, R = red, FR = far red) indicates its photon flux density in µmol•m–2•s–1. Seedling heights followed by the same letter are statistically similar.
In addition, at the end of the plug stage, seedlings were transplanted into 4-in. pots and subsequently grown in a common greenhouse at 68F (20C) with a 16-hour photoperiod to determine whether the lighting treatments had any residual effects after transplant.
For geranium and petunia, days to the first open flower from transplant, the number of visible flower buds or inflorescences, and plant height at flowering were evaluated. For coleus, plant height was evaluated when they were considered of marketable size. The experiment was performed twice and data were statistically analyzed.
Research results
Stem elongation and leaf expansion. At both PPFDs, seedling height of all species increased as far-red light was added (Figure 1). The addition of far red also increased the total leaf area of petunia at both PPFDs, while coleus showed similar trends only under the low PPFD. Generally, stem elongation and leaf expansion increased linearly as the portion of the added far-red light increased. In geranium, lighting treatments had little or no effect on total leaf area.
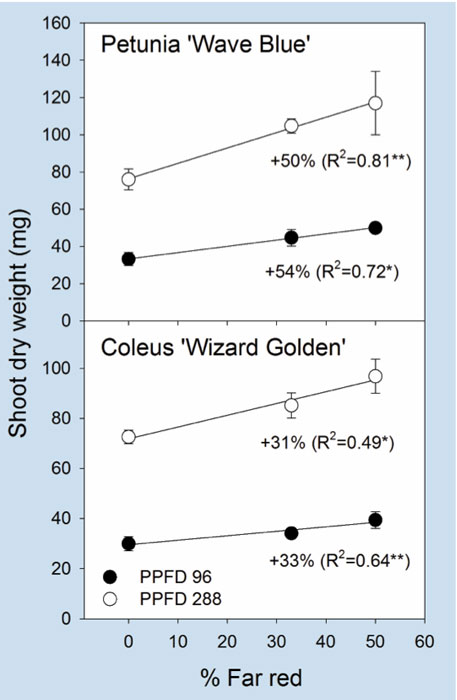 Figure 2. Influence of adding far-red light on shoot dry weight of petunia and coleus grown under a photo-synthetic photon flux density (PPFD) of 96 or 288 µmol•m–2•s–1 (see Table 1 for details). The percentage of far red was calculated as the photon flux density of far red divided by the sum of red and far-red photon flux density. Each data point represents the mean and standard error. Associated correlation coefficients (R2) and the percentage increase are presented when statistically significant. * or ** indicate significant at P<0.05 or 0.01, respectively.
Figure 2. Influence of adding far-red light on shoot dry weight of petunia and coleus grown under a photo-synthetic photon flux density (PPFD) of 96 or 288 µmol•m–2•s–1 (see Table 1 for details). The percentage of far red was calculated as the photon flux density of far red divided by the sum of red and far-red photon flux density. Each data point represents the mean and standard error. Associated correlation coefficients (R2) and the percentage increase are presented when statistically significant. * or ** indicate significant at P<0.05 or 0.01, respectively.
Dry weight. Shoot dry weight is a good indicator of plant growth. It increased linearly by up to 54% (in petunia) and 33% (in coleus) as the portion of far-red light increased at both PPFDs (Figure 2). In petunia, shoot dry weight increased linearly with total leaf area at both PPFDs, indicating that inclusion of far-red light can increase plant growth, at least partly by increasing leaf size. There was little or no effect of far-red light on shoot dry weight of geranium and root growth of any species.
Subsequent flowering and plant height at finishing. In the long-day plant petunia, the addition of far red during the seedling stage accelerated subsequent flowering at both PPFDs (Figure 3). At each PPFD, flowering was promoted similarly when the R:FR was 2:1 or lower. The addition of far-red light under PPFD 96 accelerated flowering by 11 days, whereas it only accelerated flowering by seven days at PPFD 288. In day-neutral geranium, the addition of far red had no effect on flowering at each PPFD. However, the plants grown under the higher PPFD with far-red light flowered nine or 10 days earlier than plants grown under the lower PPFD without
far-red light.
Figure 3. Influence of adding far-red light during the seedling stage on days from transplant to flowering of petunia and geranium. The subscript value after each LED type (B = blue, R = red, FR = far red) indicates its photon flux density in µmol•m–2•s–1. Flowering times followed by the same letter are statistically similar.
The li ghting treatments had no effect on stem length at flowering in geranium and petunia. Similarly, stem length of coleus at the finishing stage was similar, regardless of lighting treatments during the seedling stage (Figure 4).
ghting treatments had no effect on stem length at flowering in geranium and petunia. Similarly, stem length of coleus at the finishing stage was similar, regardless of lighting treatments during the seedling stage (Figure 4).
Conclusions
Light intensity and quality interact to regulate plant growth and development, and thus, the spectral effects of specific wavelengths can vary depending on the spectral and intensity combination. Here, we investigated the effects of adding far-red light in indoor LED lighting on ornamental seedling growth and subsequent flowering under two light intensities. Our results show how the inclusion of far-red light can promote seedling growth, regardless of light intensity, when blue light  intensity is kept constant. However, far red and light intensity interact to regulate the subsequent flowering.
intensity is kept constant. However, far red and light intensity interact to regulate the subsequent flowering.
Figure 4. Influence of adding far-red light during the seedling stage on plant height (in cm) of coleus at the finishing stage. The subscript value after each LED type (B = blue, R = red, FR = far red) indicates its photon flux density in µmol•m–2•s–1. Plant heights followed by the same letter are statistically similar.
In long-day petunia, far red had a greater effect on accelerating subsequent flowering under the lower light intensity than under higher light intensity. In day-neutral geranium, far-red light promoted subsequent flowering only when it was provided with the higher light intensity. GT
Acknowledgements: We thank the American Floral Endowment, OSRAM Opto Semiconductors, OSRAM Innovation, MSU’s AgBioResearch, and Floriculture Research Alliance partners and horticultural companies for supporting research and outreach programs at MSU.
Yujin Park (parkyuj2@msu.edu) is a Post-doctoral Research Associate and Erik Runkle (runkleer@msu.edu) is Professor and Floriculture Extension Specialist in the Department of Horticulture at Michigan State University.